V(D)J rekombinasyonu
| |
Bu sayfanın veya bölümün Türkçeye çevrilmesi gerekmektedir. Eğer sayfanın Türkçeye çevrilmesi en fazla 2 hafta içinde gerçekleşmezse, bu sayfa veya bölüm silinme sürecine girecektir. |
V(D)J rekombinasyonu (somatik rekombinasyon olarak da bilinir), bağışıklık sisteminin immunoglobulin (Ig) ve T hücre reseptörlerinin (TCR) üretiminin erken evrelerindeki genetik rekombinasyon mekanizmalarıdır.
V(D)J rekombinasyonu birincil lenfoid organlarda yani; B hücreleri için kemik iliği ve T hücreleri için timüsda meydana gelir.
V(D)J rekombinasyonu; omurgalıların V : değişken (variable) D : çeşitli (diverse) J : katılım (joining) gen kısımlarını hemen hemen rastgele birleştirir, ve farklı genlerin seçilmesindeki bu rastgelelik yüzünden, bakteri, virüs, parazitler ve işlevsiz tümör hücreleri [1] ve polenlerden gelen antijenlerle eşleşecek olan proteinlerin farklı şekillerde kodlanmaları mümkün olmaktadır.
Arka plan
İnsan antikor molekülleri (B hücre reseptörleri), üç lokustaki genler tarafından kodlanan C (daimi) ve V (değişken) bölgelerinin her ikisini de içeren ağır ve hafif zincirlerden oluşurlar.
- İmmunoglobulin ağır lokus (IGH@) 14. kromozom üzerindedir ve immunoglobulin ağır zincir için genleri içerir.
- İmmunoglobulin kappa (κ) lokus (IGK@) 2. kromozom üzerindedir ve immunoglobulin hafif zincir için genleri içerir.
- İmmunoglobulin lambda (λ) lokus (IGL@) 22. kromozom üzerindedir ve immunoglobulin hafif zincir için genleri içerir.
İnsan genomunda kodlanan değişken bölgeler için kodlanan çoklu genler bileşenlerin üç farklı tipini içerirler. Örneğin immunoglobulin ağır zincir 44 değişken (V) gen bölgesi [2] 27'den fazla Çeşitlilik (D) geni ve 6 birleşme (J) geni içermektedir.[3] Hafif zincir ayrıca bir hayli çok sayıda V be J genlerine sahiptir ancak D geni içermez.
Bu bölgesel genlerin DNA tekrar-düzenlemelerindeki mekanizmayla, muazzam büyüklükte antikor dağarcığını oluşturmak mümkündür; bazıları kendinden-olan bileşenlere bağlanabildiği için ortadan kaldırılıyor olsa da; kabaca 3×1011 kombinasyon oluşturulabilir.
T hücre reseptörlerinin çoğu bir alfa ve bir beta zincirden oluşurlar. T hücre reseptörü genleri immünoglobulin genlerine, lenfosit gelişimi sırasında eşsiz bir antijen reseptörü sağlayan, beta zincirlerinde çoklu V, D ve J ve alfa zincirde V ve J gen bölgeleri içerdiklerinden dolayı benzerlik gösterirler. Hücrenin süreci oluştururken yaptığı herhangi bir hata, hücrenin kendine tepki vermemesi için apoptozla sonuçlanır.
İmmunoglobulinlerde
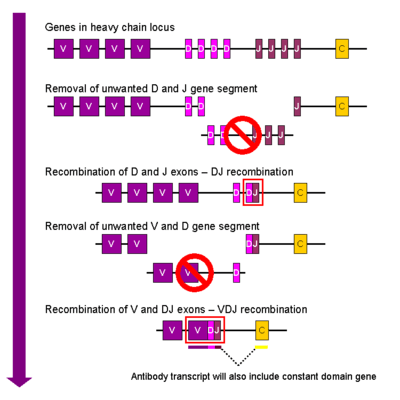
Ağır zincir
B hücrelerinin gelişmesinde, ilk rekombinasyon durumu, ağır zincir lokusunun bir D ve bir J gen bölümünde meydana gelir. Bu iki gen arasındaki herhangi bir DNA kısmı silinir. Bu D-J rekombinasyonu yeni oluşan DJ kompleksinin üstksımında bir bölgeye bir V geninin katılmasıyla ve düzenlenmiş yeni VDJ geninin oluşumuyla tamamlanır. Yeni VDJ geninin V ve D parçaları arasındaki tüm genler hücrenin genomundan artık silinebilir.
Primary transcript (unspliced RNA) is generated containing the VDJ region of the heavy chain and both the constant
Ana transkript (splayslanmamış RNA) ağır zincirin VDJ bölgesini içeren mu ve delta zincirlerinin her ikisi de değişmezdir (Cμ and Cδ). (örn. ana trankript şu segmentleri içerir: V-D-J-Cμ-Cδ). Ana transkriptin oluşturma süreci, bir poli A kuyruğunun Cμ zincirinden sonra eklenmesini ve VDJ bölgesi arasındaki bölgenin ve bu değişmez gen bölgesinin oluşturulmasını içerir. Bu mRNA'nın translasyonu Ig μ ağır zincir proteinin üretilmesine yol açar.
Hafif zincir
İmmünoglobulin hafif zincirinin lokusunun Kappa (κ) ve Lambda (λ) zincirleri, D bölgesine sahip olmayan bir hafif zincir farklılığı dışında, oldukça benzer bir süreçle üretilir. Başka bir deyişle, hafif zincirlerin rekombinasyonu için ilk basamak, birinci transkripsiyon esnasında V ve J zincirlerinin VJ kompleksi oluşturmak üzere değişmez diziden önce eklenmesidir.
Kappa ya da lambda zincirlerin her ikisi için de, splayslanmış mRNA'nın translasyonu, Ig κ ya da Ig λ hafif zincir proteinlerinin oluşumuyla sonuçlanır. Ig μ ağır zincirinin ve bu hafif zincirlerden birinin kuruluşunda, olgun B hücresinin yüzeyinde ifade edilen membrana bağlı immünoglobulin IgMnın oluşumu görülür.
In T cell receptors
During thymocyte development, the T cell receptor (TCR) chains undergo essentially the same sequence of ordered recombination events as that described for immunoglobulins. D-to-J recombination occurs first in the β chain of the TCR. This process can involve either the joining of the Dβ1 gene segment to one of six Jβ1 segments or the joining of the Dβ2 gene segment to one of seven Jβ2 segments. DJ recombination is followed (as above) with Vβ-to-DβJβ rearrangements. All genes between the Vβ-Dβ-Jβ genes in the newly formed complex are deleted and the primary transcript is synthesized that incorporates the constant domain gene (Vβ-Dβ-Jβ-Cβ). mRNA transcription splices out any intervening sequence and allows translation of the full length protein for the TCR Cβ chain.
The rearrangement of the alpha (α) chain of the TCR follows β chain rearrangement, and resembles V-to-J rearrangement described for Ig light chains (see above). The assembly of the β- and α- chains results in formation of the αβ-TCR that is expressed on a majority of T cells.
Mechanism
Recombination signal sequences
The regional genes (V, D, J) are flanked by Recombination Signal Sequences (RSSs) that are recognized by a group of enzymes known collectively as the VDJ recombinase. RSSs are composed of seven conserved nucleotides (a heptamer) that reside next to the gene encoding sequence followed by a spacer (containing either 12 or 23 unconserved nucleotides) followed by a conserved nonamer (9 base pairs). The RSSs are present on the 3’ side (downstream) of a V region and the 5’ side (upstream) of the J region. These are the sides that will be involved in the joining. Only a pair of dissimilar spacer RSSs are efficiently recombined (i.e. one with a spacer of 12 nucleotides will be recombined with one that has a spacer containing 23 nucleotides). This is known as the 12/23 rule of recombination (or "one turn/two turn" rule).
VDJ recombinase
VDJ recombinase refers to a collection of enzymes some of which are lymphocyte specific, and some that are expressed in many cell types. The initial steps of VDJ recombination are carried out by critical lymphocyte specific enzymes, called recombination activating gene-1 and -2 (RAG1 and RAG2). These enzymes associate with each other to recognize the RSS sequences and induce DNA cleavage at the RSS sequences. This cleavage only takes place on one strand of DNA, which leads to a nucleotide attack and creation of a hairpin loop.
Other enzymes of the VDJ recombinase are expressed in multiple cell types and are involved in DNA repair following the activity of RAG1 and RAG2. RAG1 is homologous to transposase.[4] One of these enzymes is called the DNA-dependent protein kinase complex (DNA-PK) that repairs double-stranded DNA. DNA-PK binds to each end of the broken DNA and recruits several other proteins, including Artemis nuclease, XRCC4 (X-ray repair cross-complementing factor 4), DNA ligase IV, Cernunnos (also called XLF or XRCC4-like factor), and any of several DNA polymerases. DNA-PK complexes on each DNA end phosphorylate (add phosphate groups to) each other, resulting in activation of Artemis. Artemis then breaks the hairpin loop that was formed by the RAG proteins.[5] XRCC4 and Cernunnos act in concert with DNA-PK to align the two DNA ends with each other, and also help to recruit the enzyme, terminal deoxynucleotidyl transferase (TdT), which adds nucleotides randomly to the ends, providing junctional diversity. DNA polymerases λ and μ insert additional nucleotides as needed to make the two ends compatible for joining. Ligase IV finally links DNA strands on opposite ends of the break to each other, completing the joining process.[6]
Because of the variability in the exact position of cleavage of the hairpin loop by Artemis, as well as the random nucleotide addition by terminal deoxynucleotidyl transferase (TdT), the final DNA sequence, and thus the sequence of the resulting antibody, is highly variable, even when the same two V, D, or J segments are joined. This great diversity allows VDJ recombination to generate antibodies even to microbes that neither the organism nor its ancestors have ever previously encountered.
Kaynakça
- ↑ Abbas AK and Lichtman AH (2003). Cellular and Molecular Immunology (5th bas.). Saunders, Philadelphia. ISBN 0-7216-0008-5.
- ↑ Matsuda, F; Ishii, K; Bourvagnet, P; Kuma, K; Hayashida, H; Miyata, T; Honjo, T (1998). "The complete nucleotide sequence of the human immunoglobulin heavy chain variable region locus". The Journal of experimental medicine 188 (11): 2151–62. DOI:10.1084/jem.188.11.2151. PMC 2212390. PMID 9841928. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=2212390.
- ↑ Li A, Rue M, Zhou J ve diğ. (June 2004). "Utilization of Ig heavy chain variable, diversity, and joining gene segments in children with B-lineage acute lymphoblastic leukemia: implications for the mechanisms of VDJ recombination and for pathogenesis". Blood 103 (12): 4602–9. DOI:10.1182/blood-2003-11-3857. PMID 15010366. http://www.bloodjournal.org/cgi/pmidlookup?view=long&pmid=15010366.
- ↑ V.V. Kapitonov and J. Jurka (2005). "RAG1 core and V(D)J recombination signal sequences were derived from Transib transposons". PLoS Biol. 3 (6): e181. DOI:10.1371/journal.pbio.0030181. PMC 1131882. PMID 15898832. http://www.pubmedcentral.nih.gov/articlerender.fcgi?tool=pmcentrez&artid=1131882.
- ↑ Y. Ma, U. Pannicke, K. Schwarz and M.R. Lieber (2004). "Hairpin opening and overhang processing by an Artemis/DNA-dependent protein kinase complex in nonhomologous end joining and V(D)J recombination". Cell 108 (6): 781–794. DOI:10.1016/S0092-8674(02)00671-2. PMID 11955432.
- ↑ D.C. van Gent and M. van der Burg (2007). "Non-homologous end-joining, a sticky affair". Oncogene 26 (56): 7731–7740. DOI:10.1038/sj.onc.1210871. PMID 18066085.
Ek okuma
- Hartwell LH, Hood L, Goldberg ML, Reynolds AE, Silver LM, Veres RC (2000). Chapter 24, Evolution at the molecular level. In: Genetics. New York: McGraw-Hill. s. 805–807. ISBN 0-07-299587-4.
- V(D)J Recombination. Series: Advances in Experimental Medicine and Biology, Vol. 650 Ferrier, Pierre (Ed.) Landes Bioscience 2009, XII, 199 p. ISBN 978-1-4419-0295-5