Genetik
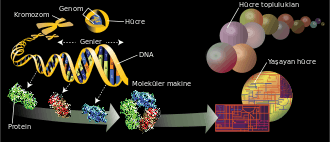
Genetik ya da kalıtım bilimi[1], biyolojinin organizmalardaki kalıtım ve çeşitliliği inceleyen bir dalı.[2][3] Türkçeye Almancadan geçen[1] genetik sözcüğü 1831 yılında Yunanca γενετικός - genetikos ("genitif") sözcüğünden türetildi. Bu sözcüğün kökeni ise γένεσις - genesis ("köken") sözcüğüne dayanır.[4]
Canlıların özelliklerinin kalıtsal olduğunun bilinci ile, tarih öncesi çağlardan beri bitki ve hayvanlar ıslah edilmiştir. Bununla birlikte, kalıtımsal aktarım mekanizmalarını anlamaya çalışan modern genetik bilimi ancak 19. yüzyılın ortalarında, Gregor Mendel’in çalışmasıyla başlamıştır.[5] Mendel, kalıtımın fiziksel temelini bilemediyse de, bu özelliklerin ayrık (kesikli) bir tarzda aktarıldığını gözlemlemiştir; günümüzde bu kalıtım birimlerine "gen" adı verilmektedir.
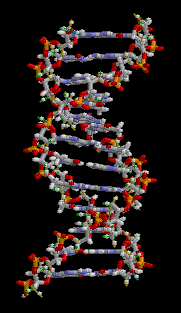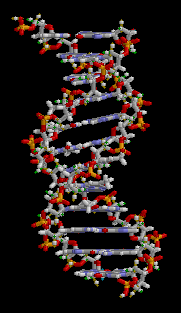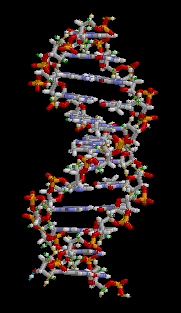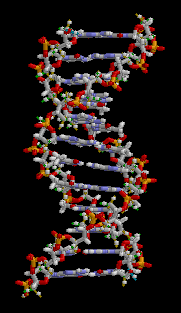
Genler DNA'da belli bölgelere karşılık gelir. DNA dört tip nükleotitten oluşan bir zincir moleküldür. Bu zincir üzerinde nükleotitlerin dizisi, organizmaların kalıt aldığı genetik bilgidir (enformasyon). Doğada DNA, iki zincirli bir yapıya sahiptir. DNA'daki her "iplikçik"teki nükleotitler birbirini tamamlar, yani her iplikçik, kendine eş yeni bir iplikçik oluşturmak için bir kalıp olabilme özelliğine sahiptir. Bu, genetik bilginin kopyalanması ve kalıtımı için işleyen fiziksel mekanizmadır.
Nükleotitlerin DNA’daki dizilişi, hücre tarafından amino asit zincirleri üretmek için kullanılır. Bunlardan protein oluşur. Bir proteindeki amino asitlerin sırası, gendeki nükleotitlerin sırasına karşılık gelir. Aradaki bu ilişkiye genetik kod denir. Amino asitlerin bir proteindeki dizilişi, proteinin nasıl bir üç boyutlu şekil alacağını belirler. Bu yapının şekli de proteinin fonksiyonundan sorumludur. Hücrelerin yaşamaları ve üremeleri için gerekli hemen hemen tüm fonksiyonları proteinler icra ederler. DNA dizisindeki bir değişim, bir proteinin amino asit dizisini ve dolayısıyla onun şekli ve fonksiyonunu değiştirir: Bu, hücrede ve onun bağlı bulunduğu canlıda önemli sonuçlara yol açabilir.
Genetik, organizmaların görünüşünün ve davranışının belirlenmesinde önemli bir rol oynuyorsa da, sonucun oluşmasında, organizmanın çevre ile etkileşimi ve genetik birlikte etki eder. Örneğin genler kişinin boyunun uzunluğunda bir rol oynuyorsa da, kişinin çocukluk çağındaki beslenmesinin ve sağlığının da büyük bir etkisi vardır.
Tarihçe
- Konu hakkında ayrıntılı bilgi için Genetik tarihi maddesine bakınız.
Genetik bilimi 1800'lü yılların ortalarında Gregor Mendel'in uygulamalı ve teorik çalışmalarıyla başladıysa da, kalıtım ile ilgili başka teoriler Mendel'den önce mevcuttu. Mendel'in zamanında popüler olan bir teori, karışmalı kalıtım kavramıydı: Bireylerin, ebeveyninin özelliklerinin homojen bir karışımını kalıt aldığı fikriydi bu. Mendel'in çalışmaları bunu yanlışladı, özelliklerin ayrık genlerin birleşimi olduğunu, sürekli özelliklerin bir karışımı olmadığını gösterdi. (Örneğin, kırmızı ve beyaz gözlü sinekler çiftleştiğinde yavruları ya kırmızı ya beyaz gözlü olur, ama pembe gözlü olmaz.) O devirde geçerli olan bir diğer teori, edinilmiş özelliklerin kalıtımı idi: kişilerin ebeveyninin kuvvetlendirdiği özellikleri taşıdığı inancıydı. Bu fikrin (genelde Jean-Baptiste Lamarck'a atfedilir) bugün yanlış olduğu bilinmektedir.
Kişilerin deneyimleri, yavrularına aktardıkları genleri değiştirmez. Diğer teoriler arasında Charles Darwin'in Pangenezis fikri (ki bu hem kalıtsal hem de edinilmiş özellikler öne sürer) ve Francis Galton'un Pangenezis'e getirdiği yeni bir yorum olarak, kalıtımın hem tanecikli hem de kalıtsal olduğu fikriydi.
İlk genetik deneyi, Mendel ve Klasik Genetik

Modern genetik biliminin kökü, Avusturyalı (Alman-Çek) bir Augustin’ci[6] keşiş ve bir botanikçi olan Gregor Johann Mendel’in gözlemlerine dayanır.
Günümüzün bu popüler biliminin babası olarak kabul edilen Mendel, bitkilerde kalıtım özellikleri üzerine ayrıntılı çalışmalar yapmıştır. Mendel 1856 yılından itibaren çeşitli bezelye (Pisum sativum) varyetelerine ait tohumları toplamaya ve onları manastır bahçesinde yetiştirerek aralarındaki farkları incelemeye başladı. 10 yıl süren gözlem ve deneylerinin ardından, bu çalışmasının önemli bulgularını “Versuche Über Pflanzenhybriden” (“Bitki melezleri üzerinde denemeler”) adlı ünlü inceleme yazısıyla yayımladı ve bu yazıyı 1865’te Brunn Doğa Tarihi Derneğine sundu. Mendel, bezelye bitkilerindeki bazı özelliklerin kalıtımsal tekrarını izlemiş ve bunların matematiksel olarak tanımlanabileceğini göstermiştir.[7] Mendel'in çalışması kalıtımın edinilmiş değil, tanecikli olduğunu ve pek çok özelliğin kalıtımının basit kural ve orantılar ile açıklanabileceğini öne sürmüştür.
O tarihlerde DNA, kromozom, mayoz bölünme gibi kavramların henüz ortaya konmamış olduğu ve bilinmediği göz önüne alınırsa, Mendel’in sadece fenotipik (gözlenebilen) karakter ayrılıklarına göre yapmış olduğu değerlendirmelerin son derece başarılı olduğu söylenebilir.
Mendel'in ölümünden sonra gelen 1890'lara kadar, onun çalışmasının önemi geniş çaplı olarak anlaşılamadı. O dönemde benzer problemler üzerinde çalışan başka bilimciler onun çalışmalarını tekrar keşfettiler. Ölümünden 16 yıl sonra Hollanda’da Hugo De Vries, Almanya’da Correns ve Avusturya’da E. Von Tschermak adlı üç biyolog, çeşitli bitki türlerinde, birbirlerinden habersiz yaptıkları araştırmalarda, Mendel yasalarının geçerliliğini gösterdiler ve tüm sonuçları "Mendel yasaları" adı altında toparladılar. Mendel'in çalışması aynı zamanda, kalıtım çalışmalarında istatistik yönteminin kullanımını önermekteydi.[8]
"Genetik" terimi, 1905’te Mendel’in çalışmasının önemli savunucularından William Bateson tarafından Adam Sedgwick’e gönderilen bir mektupta ortaya atılmıştır.[9][10] Bateson 1906’da Londra’da yapılan Üçüncü Uluslararası Bitki Melezleri Konferansı’nda yaptığı açış konuşmasında kalıtım çalışmasını tanımlarken “genetik” terimini kullanarak, bu terimin yaygınlaşmasını sağlamıştır.[11] (bir sıfat olarak genetik, Yunanca genesis - γένεσις ("kaynak")'tan türemiştir, o da genno - γεννώ ("doğurmak")'tan; biyolojik anlamıyla bu sıfat, isim hâliyle 'genetik'ten daha önce, ilk defa 1860'ta kullanılmıştır)[12])
Mendel’in çalışmasının yeniden keşfinin ve popüler hâle gelişinin ardından, DNA moleküler temelini gün ışığına çıkarmaya yönelik birçok deney yapılmıştır. Beyaz gözlü Drosophila (meyve sineği) üzerindeki gözlemlerinden yola çıkan Thomas Hunt Morgan 1910’da genlerin kromozomlarda yer aldığını ileri sürmüş ve 1911’de mutasyonların varlığını ortaya koymuştur.[13] Morgan'ın öğrencisi Alfred Sturtevant ise genetik bağlantı fenomenini kullanmış ve 1913’te genlerin kromozom boyunca birbirini izleyen dizilişi ve düzenini gösteren, ilk “genetik harita”yı yayımlamıştır.[14]
Moleküler genetik
Önceleri, kromozomların genleri içerdiği ve protein ile DNA’dan oluştuğu bilinmekteyse de, kalıtımdan hangisinin sorumlu olduğu bilinmiyordu. 1928’de Frederick Griffith, yayımladığı makalesinde, keşfettiği transformasyon fenomenini açıkladı. Bundan 16 yıl sonra da, 1944'te, Oswald Theodore Avery, Colin McLeod ve Maclyn McCarty bu transformasyondan sorumlu molekülün DNA olduğunu gösterdiler.[15] 1952'deki Hershey-Chase deneyi de, DNA'nın (proteinden farklı olarak) virüslerin genetik malzemesi olduğunu, diğer molekülün kalıtımdan sorumlu olamayacağını kanıtladı.[16]
James D. Watson ve Francis Crick 1953'te DNA'nın yapısını çözdüler ve Rosalind Franklin'in çalışması olan X ışını kırınım çalışması sonuçlarını kullanarak DNA molekülünün sarmal bir yapısı olduğunu gösterdiler.[17][18] Onların ikili sarmal modeli, nükleotit dizisinin diğer iplikçikte tamamlayıcı eşleri olduğunu gösterdi.[19] Bu yapı, nükleotitlerin sıralanmalarıyla genetik bilginin saklanabileceğini göstermekle kalmadı, aynı zamanda ikileşme için fiziksel mekanizmasını gösterdi: iki iplikçik birbirinden ayrışınca, her iplikçik kendine eş olacak yeni bir iplikçiğin oluşumu için kendi dizisini bir kalıp olarak kullanabilirdi.
Bu yapı, kalıtım sürecini açıklamaktaysa da; DNA’nın hücre davranışlarını nasıl etkilediği henüz bilinmiyordu. Sonraki yıllarda, bazı bilim insanları, DNA'nın, ribozomlardaki protein üretim süreçlerini kontrol mekanizmasını anlamaya çalıştılar ve DNA'nın genetik kodunun mesajcı RNA (mRNA) ile okunduğunu ve çözüldüğünü buldular.[20] RNA, DNA'ya benzer, nükleotitlerden oluşmuş bir moleküldür; mRNA'nın nükleotit dizisi proteinlerdeki amino asit dizisini oluşturmak için kullanılır. Nükleotit dizisinin amino asit dizisine çevirisi genetik kod aracılığıyla gerçekleşir.
Kalıtım konusunda yapılan bu moleküler düzeydeki buluşlar, DNA'nın moleküler yapısının anlaşılmasını ve biyolojideki yeni bilgilere uygulanan bir araştırma patlamasını sağlamıştı. 1977’de Frederick Sanger'in zincir sonlandırmalı DNA dizileme yöntemi önemli bir gelişme olmuştur; bu teknoloji bilimcilerin DNA moleküllerini okumasını sağlamıştır.[21] 1983'de Kary Mullis tarafından geliştirilen polimeraz zincir tepkimesi ise, DNA izolasyonunu ve DNA parçalarının istenen bölgelerinin kolayca çoğaltılmasını sağladı.[22] Bu ve diğer teknikler ve bir yandan İnsan Genom Projesi’nin ekip çalışması, diğer yandan Celera Genomics’in özel çalışması sonucunda, 2003’te insan genomu dizileri tümüyle gün ışığına çıkarılmıştır.[23]
Kalıtım özellikleri
Kesikli kalıtım ve Mendel yasaları
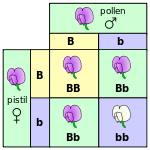
En temel düzeyde, organizmalardaki kalıtım, günümüzde genler adını verdiğimiz ayrık özellikler aracılığıyla meydana gelir.[24] (Bir özelliğin büyüklüğü iki, veya birkaç değer etrafında toplanmışsa bu özellik ayrıktır; eğer sürekli bir değerler dağılımı gösteriyorsa, süreklidir) Bu konuda gözlemde bulunan ilk kişi, bezelye bitkisi de kalıtımsal özelliklerinin ayrışımı üzerinde çalışmış Gregor Mendel olmuştur.[25][26] Çiçek rengi üzerine yaptığı araştırmalarda, Mendel her bir çiçeğin ya mor ya beyaz olduğunu, ara bir renk olmadığını gözlemledi. Aynı genin farklı, birbirinden ayrık versiyonları alel olarak adlandırılır.
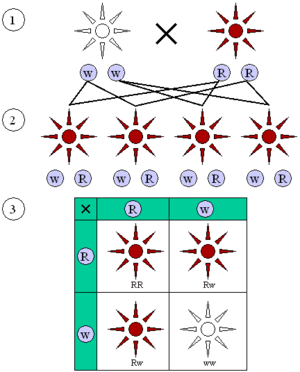
Mendel farklı bitki çeşitlerinin her birinden tohumlar toplayarak bahçesinde ekti. Bezelye bitkilerini düzenli “tozlaşma”lara tabi tutan Mendel, bunlarda 7 özelliğin değişmediğini keşfetti ve bezelyelerdeki bu 7 özelliğin (tanelerin biçimi, rengi, bitkilerin boyu vs.) dölden döle nasıl aktarıldığını gözlemledi. Her dölde elde ettiği bireyleri, birbirlerine ve ebeveynine benzeyip benzemediklerine göre ayrıma tâbi tuttu. Böylece özellikleri farklı 7 saf döl elde etti. Bunlarla yaptığı çaprazlamalarda[27] bazı belirli özelliklerin değişmediğini saptadı. Bu özelliklerin her birine “saf özellik” adını verdi. İki eş "saf özellik" çaprazlandığında, sadece bu saf özellik ortaya çıkmaktaydı ki, Mendel yasalarının esasını teşkil eden de bu husustur.

Mendel, ayrıca, yaptığı çaprazlamalarda bazı özelliklerin baskın olduğunu gözlemledi. Örneğin, uzunluk karakteri, kısalık karakterine baskın olduğundan, melez bireyler uzun görünümdeydi. İki uzun melezin çaprazlanması sonucunda ise % 25 oranında saf uzun, % 25 saf kısa, % 50 melez uzun çıkmaktaydı.
Mendel, bezelye bitkisinin çiçeklerinin rengi üzerindeki deneme çalışmasında, rengin ya mor ya da beyaz olduğunu ve asla bu iki rengin karışımı bir rengin oluşmadığını gözlemledi. Aynı genin bu farklı versiyonlarına alel adı verilir.
Bezelye bitkilerinde her organizma her genin iki aleline sahiptir.[28] İnsan da dahil olmak üzere birçok organizmada bu kalıtım modeli geçerlidir. (Genetikte böyle bir organizmadaki genin iki alelinden birinin anneden, diğerinin babadan geçtiği kabul edilir.) Aynı alelin iki kopyasını içeren organizmalara homozigot, iki farklı alele sahip organizmalara ise heterozigot adı verilir.
Bir organizmadaki alellerden oluşan genetik yapısına genotip denir. Organizmanın sahip olduğu gözlemlenebilir özelliklere ise fenotip adı verilir.
Heterozigot organizmalarda genellikle, alellerden birinin nitelikleri diğerininkileri bastıracak şekilde organizmanın fenotipini belirler; alellerden nitelikleri organizmanın fenotipine hakim olanına (baskın çıkana) "baskın" (dominant), niteliklerinin fenotipe hakim olmadığı gözlemlenen öteki alele ise "çekinik" (resesif) adı verilir. Bununla birlikte, bazen bir alelin tam anlamıyla baskın olmadığı görülmüştür ki, bu duruma “eksik baskınlık” adı verilir. Bazen de her iki alelin niteliklerinin birden etkili olduğu gözlemlenir ki, bu duruma da “eşbaskınlık” (kodominans) adı verilir.[29]
Bir çift organizma çiftleştiğinde, döl (yavru), rastgele bir biçimde, iki alelinden birini anneden, diğerini babadan miras (kalıt) alır. Ayrık kalıtım ve alellerin ayrışımı üzerine yapılmış bütün bu gözlemler, toplu olarak, Mendel’in birinci yasası veya Ayrışma Yasası adıyla bilinir.

Sembolik gösterim sistemi ve şemalar
Genetikçiler kalıtımı betimlemede şema ve semboller kullanırlar. Bir gen bir veya birkaç harfle temsil edilir. Bu temsilde büyük harf baskın aleli, küçük harf çekinik aleli temsil eder.[30] Genellikle bir “+” sembolü bir gen için normal, mutant olmayan aleli temsil etmede kullanılır. Döllenmede ve Mendel’le ilgili üretme deneylerinde ebeveyn, "parent" sözcüğünün başharfi olan “P” ile, döl (yavru) F1 ile (“F” "filial" sözcüğünün başharfi, “1” de birinci nesil anlamında) ile değinilir. F1 neslindeki döller birbiriyle çiftleşince meydan gelen yeni nesildeki döller F2 olarak değinilir. Çaprazlamanın sonucunu öngörmede kullanılan yaygın şemalardan biri "Punnett karesi" olarak bilinir.
Genetikçiler insandaki genetik hastalıkları incelerken genellikle, özelliklerin kalıtımını temsil etmede soyağacı çizelgesi kullanırlar.[31]
Genlerin etkileşimi

Organizmalar binlerce gen içerir ve cinsel çiftleşmeyle üreyen organizmalarda bu genlerin birlikte bulunmaları (tertiplenmeleri) genellikle birbirlerinden bağımsızdır. Yani, örneğin, sarı veya yeşil renkli bir bezelye alelinin kalıtımı (aktarımı), çiçeklerin beyaz veya mor oluşunu belirleyen alellerin kalıtımıyla ilişkisizdir. “Mendelin ikinci yasası” veya “Bağımsız Tertiplenme Yasası” olarak bilinen bu olguda, ebeveynin her ikisinden gelerek karışan farklı genlerin alellerinin, dölü oluştururken farklı pek çok kombinasyonla bir araya gelebileceği anlamına gelir. (Ancak "Genetik bağlantı" gösteren bazı genler bağımsız olarak bir araya gelmezler edilmezler, bu konu aşağıda daha ayrıntılı işlenecektir.)
Sıkça görüldüğü gibi, farklı genler aynı özelliği (fenotipi) oluşmasını sağlayacak tarzda birbirlerini etkileyebilirler. Avrupa kökenli Omphalodes verna bitkisinin genleri bu duruma örnek olarak gösterilebilir. Bu bitkide, çiçeklerin renginin mavi ya da magenta olmasını sağlayan iki alelli bir gen vardır. Fakat bu bitkide bir de, çiçeklerin renkli olup olmayacağını, yani renkli veya beyaz olacağını denetleyen, iki alelli bir başka gen daha vardır. Bitki bu ikinci genin beyaz alelinin iki kopyasına sahip olduğu zaman, birinci gendeki mavi ile magenta rengi alellerden birinin bitkide etkili olmasına meydan verilmeksizin, çiçekler beyaz olur. Genler arasındaki bu etkileşime "epistasis" adı verilir, sıfat olarak da, birinci genin ikincisi üzerinde "episatik" olduğu söylenir.[32]
Birçok özellik ayrık özellik (beyaz ya da mor çiçekler örneğinde olduğu gibi) olmak yerine sürekli özelliktir (insan boyu ve deri rengi gibi). Bu karmaşık özellikler birçok genin ürünüdür.[33] Bu genlerin etkisi, organizmanın deneyimlerde bulunduğu çevrenin etkileriyle çeşitli derecelerde dengelenir. Bir organizmanın genlerinin böyle bir karmaşık özelliğe katkıda bulunma derecesine “kalıtsallık” adı verilir.[34] Bir özelliğin kalıtsallık ölçüsü, çevrenin o özellik üzerindeki değişen etkilerine bağlı olarak görecelidir. Örneğin insanın boyu dediğimiz karma özelliğin kalıtsallığı A.B.D.’nde %89 olarak belirlenmişken, beslenme ve sağlık sorunlarının bulunduğu Nijerya gibi yoksul bir ülkede çevrenin etkisi daha büyük olduğundan, bu oran ancak %62 olarak belirlenmiştir.[35]
Kalıtımın moleküler temeli
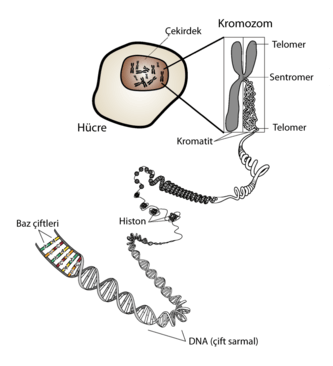
DNA ve kromozomlar
Genlerin moleküler temeli deoksiribonükleik asittir (DNA). DNA da 4 tipteki bir nükleotitler zincirinden oluşur: adenin (A), sitozin (C), guanin (G), ve timin (T). Genetik enformasyon (kalıtım bilgisi) nükleotitlerin dizilişinde bulunmakta olup, genler DNA zinciri boyunca uzanan diziler olarak mevcuttur.[36] Bu kuralın dışında kalabilen tek istisna virüslerdir; virüsler bazen DNA yerine benzeri olan RNA molekülü kullanırlar; çünkü virüslerin genetik malzemesi RNA’dır.[37]
DNA, normal olarak, ikili sarmal biçimde dolanan iki iplikçikli bir moleküldür. DNA’nın iki iplikçiğinden birindeki her nükleotit, karşıt iplikteki nükleotit partneriyle bir çift oluşturur; yani A, T ile bir çift oluşturur, C de G ile. Dolayısıyla iki iplikçikten her biri, tüm gerekli enformasyona sahip bulunur, diğer iplikçikte de bu enformasyonun yedeği bulunur. DNA’nın bu yapısı, kalıtımın fiziksel temelidir. DNA ikileşmesinde, iplikçiklerin ayrışması ve her iplikçiğin yeni iplikçik eşinin bir kalıbı olarak kullanılmasıyla, genetik enformasyon kopyalanır.[38]
Genler, kromozom denen DNA dizisi zincirleri boyunca doğrusal bir düzende sıralanmışlardır. Bakterilerde her hücrenin, basit bir dairesel kromozoma sahip olmasına karşılık, bitki ve hayvanların da dahil bulunduğu ökaryot organizmalar, çoklu doğrusal kromozomlar halinde düzenlenmiş DNA’lara sahiptirler. Bu DNA zincirleri son derece uzundur; örneğin en uzun insan kromozomu 247 milyon baz çiftini içerecek uzunluktadır.[39]
Bir kromozomdaki DNA, onu düzenleyen, sıkıştıran ve ona erişimi kontrol eden yapısal proteinlerle beraber, kromatin denen bir yapı oluşturur. Ökaryotlarda kromatin genellikle nükleozomlardan oluşur, bunlar DNA üzerinde düzenli aralıklarla yer alan, DNA'nın etrafında sarılı olduğu, histon proteinlerinden oluşmuş yapılardır.[40] Bir organizmadaki kalıtımsal malzemenin bütününe (yani, genelde, tüm kromozomlarındaki DNA dizilerinin tamamına) genom adı verilir.
Haploit organizmaların her kromozomdan yalnızca bir kopyaya sahip olmalarına karşın, hayvanların çoğu ve birçok bitkinin dahil olduğu diploitlerde, her kromozomdan iki kopya ve dolayısıyla her genden iki kopya bulunur.[28] Bir genin iki aleli, kardeş kromozomlardalerde aynı “lokus”larda (konumlarda) yer alır; bu alellerin her biri bir ebeveynden (biri anneden, biri babadan) alınmıştır.
Bunun bir istisnası, organizmanın cinsiyetinin belirlenmesinde rol oynayan, cinsiyeti belirleyen eşey kromozomlarında söz konusudur.[41] Bu kromozomlardan (örneğin insandaki 23. kromozom çiftinden), insanlarda ve memelilerde çok az gene sahip olan Y kromozomu erkeklik özelliklerinin gelişimini başlatmasına karşın, diğeri, X kromozomu, öteki kromozomlara benzemekte olup, cinsel belirlenmeyle ilgili olmayan birkaç gen içerir. Dişiler X kromozomundan iki kopyaya sahip olurlarken, erkekler bir X, bir de Y kromozomuna sahip olurlar. Dolayısıyla, cinsiyetle bağlantılı hastalıklar olarak ortaya çıkan alışılmadık kalıtım örnekleri de X kromozomunun kopyasındaki bu sayısal farklılıktan ileri gelir.
Üreme

Hücreler bölündüğünde, onların tüm genomu kopyalanır ve her yavru hücre onun bir kopyasını miras alır (kalıt alır). Mitoz adı verilen bu süreç, en sade üreme biçimi olup, “eşeysiz üreme”nin temelidir. Eşeysiz üreme, bazı çok hücreli organizmalarda da, anne veya babadan birinin genomunu miras alan bir yavru (döl) üremesini sağlayacak şekilde, oluşabilir. Genetik olarak, ebeveyninin tıpkısı olan döllere klon denir.
Ökaryotik organizmalarda ise genellikle “eşeyli üreme” olur. Eşeyli üremede ebeveynlerin her ikisinden gelen kalıtımsal materyelin karışımını içeren bir döl üretilir. Eşeyli üreme sürecinde, haploit ve diploit hücre tipleri arasında almaşık bir sıralama olur.[28] Haploit hücreler birbirleriyle kaynaşarak genetik materyelleri birleştirir ve çift kromozomlu bir diploit hücre yaratırlar. Diploit organizmalar , DNA ikileşmesi olmadan bölünerek haploit hücreler meydana getirirler. Bu yolla meydana gelen yavru haploit hücreler her kromozom çiftinden birini ya da diğerini rastlantısal olarak kalıt (miras) almışlardır. Hayvan ve bitkilerin çoğu, yaşamlarının hemen tamamını diploit olarak geçirirler, haploit biçimleri sadece, tek hücreli gametlerden ibarettir.
Bakteriler eşeyli üremenin bu haploit/diploit yöntemini kullanmasalar da, yeni kalıtımsal enformasyonun edinilmesinde birçok yöntem kullanırlar. Örneğin, bazı bakteriler konjugasyon denilen yolla, dairesel bir DNA parçasını bir bakteriden diğerine aktarırlar.[42] Bakteriler aynı zamanda, çevrelerinde bulunan DNA parçalarını alıp genomlarına dahil edebilirler ki, bu fenomen, transformasyon olarak bilinir.[43] Bu süreçler sonucunda “yatay gen aktarımı” denen, birbiriyle ilişkisiz organizmalar arasında kalıtımsal enformasyon parçalarının nakli meydana gelir.
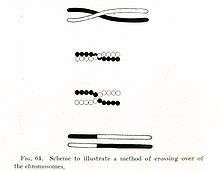
Kromozomal parça değişimi ve genetik bağlantı
Kromozomların diploit tabiatı, farklı kromozomlardaki genlere, eşeyli üreme sırasında, yeni gen kombinasyonları oluşturacak şekilde "bağımsız ayrışım" olanağı sağlar. Genlerin yeni gen kombinasyonları oluşturacak şekilde bu birleşmelerinde (rekombinasyonda), eğer kromozomların parça değiştirdiği krosover denilen süreç olmasaydı, aynı kromozomdaki genler teorik olarak asla birleşmezlerdi. Bu süreç sırasında kromozomlar, DNA parçalarını değiş tokuş yaparak, gen alellerinin değişmesini sağlarlar.[44] Bu kromozomal parça değişimi süreci genellikle mayoz bölünme sırasında, yani gametin haploit "germ hücreleri"ni yaratan bir dizi hücre bölünmesi sırasında meydana gelir. (Bu germ hücreler de daha sonra birleşerek yavru organizmayı meydana getirirler.)
Kromozomdaki belirli iki nokta arasında meydana gelebilecek rekombinasyon olasılığı bu iki nokta arasındaki uzaklığa bağlıdır. Yeterince uzak olan genler arasında hep rekombinasyon olacağından bu genlerin alleleri rastgele bir şekilde dağılırlar. Nispeten yakın genler durumunda, krosover olma olasılığının düşük olması, bu genlerin genetik bağlantı göstermesi anlamına gelir; her iki genin alelleri birlikte kalıt olmaya eğilimlidir. Genlerin dizileri arasındaki bağlantı miktarı çizgisel bir bağlantı haritası oluşturur ki, bu harita genlerin kromozom boyunca düzenlenişine kabaca karşılık gelir.[45]
Gen ifadesi
Genetik kod
Genler, fonksiyonel etkilerini, genellikle, hücredeki fonksiyonların çoğundan sorumlu, proteinlerin üretimiyle ifade ederler.[46] Proteinler amino asit zincirleridir ve bir genin DNA dizisi (bir RNA aracılığıyla) bir proteinin kendine has dizisini üretmede kullanılır. Yazılım (transkripsiyon) denilen bu süreç, genin DNA dizisine kaşılık gelen bir diziye sahip bir RNA molekülü üretimiyle başlar. Ardından, bu mesajcı RNA molekülü translasyon denilen bir süreçle, RNA dizisindeki enformasyona karşılık gelen bir amino asit dizisi üretmede kullanılır. RNA dizisindeki her üç nükleotitlik grup bir kodon olarak adlandırılır, bu kodonların her biri proteinleri oluşturan 20 amino asitten birine karşılık gelir. RNA dizisi ile amino asitler arasındaki bu ilişkiye genetik kod adı verilir.[47] Bu enformasyon akışı tek yönlü olur; yani enformasyon nükleotit dizilerinden proteinlerin amino asit dizisine aktarılır, proteinden DNA dizisine aktarılmaz. Bu olgu Francis Crick tarafından “moleküler biyolojinin merkezî dogması” olarak adlandırılmıştır.[48]

Bir proteini amino asit dizisi, o proteinin üç boyutlu yapısını oluşturur ki, bu da proteinin fonsiyonuyla yakından ilişkilidir.[49] Bunlardan bazıları, kollajen proteinince oluşturulmuş lifler gibi, basit yapılı moleküllerdir. Enzim denen proteinler başka proteinlere ve basit moleküllere bağlanabilirler, bağlandıkları moleküllerdeki kimyasal reaksiyonları kolaylaştırarak (proteinin kendi yapısını değiştirmeksizin) katalizör rolü oynarlar. Proteinin yapısı dinamiktir; örneğin hemoglobin proteini, memeli kanında oksijen moleküllerinin alınması, taşınması ve salınmasını kolaylaştırırken eğilip bükülerek farklı biçimler alır.
DNA’daki tek bir nükleotitin farkı bile, bir proteinin amino asit dizisinde bir değişikliğin olmasına neden olabilir. Proteinlerin yapıları kendi amino asit dizilerinin sonucu olduğu için de, böyle bir değişiklik o proteinin özelliklerini değiştirebilir; örneğin proteinin özelliklerini, o proteinin yapısında istikrarın bozulmasına veya o proteinin diğer protein ve moleküllerle etkileşiminde değişiklikler olmasına yol açacak şekilde, değiştirebilir. İnsanlardaki kalıtımsal hastalıklardan orak hücre anemisi adlı kan hastalığı bu duruma örnek olarak gösterilebilir. Bu hastalık, hemoglobinin β-globin bölümünü belirleyen kodlama bölgesindeki tek bir baz farklılığından kaynaklanır; bu bir bazın farklı olması, hemoglobinin fiziksel özelliklerinin değişmesine yol açan bir amino asiti değişikliğine neden olur.[50] Fiziksel özelliklerinin değişmesinin sonucunda ortaya çıkan hemoglobinin “orak hücre” versiyonları, birbirlerine yapışırlar, üstüste yığılarak lifler oluştururlar. Bu lifler proteini nakleden alyuvarların biçiminin bozulmasına yol açar. Orak biçimli hücreler kan damarları içinde rahat akamazlar, parçalanma veya damarı tıkama eğilimlidirler. Bu sorunlar sonunda kişide bu hastalıkla ilgili tıbbi rahatsızlıklara yol açar.
Bazı genler RNA’da kopyalanmakla birlikte proteine çevrilmezler ki, bunlara “kodlamayan RNA” molekülleri denir. Bu ürünler, bazı durumlarda, kritik hücre fonksiyonlar ile ilgili yapılarda rol alırlar (Ribozomal RNA, taşıyıcı RNA gibi). RNA aynı zamanda, diğer RNA molekülleriyle "hibridizasyon" etkileşimleri yoluyla düzenleyici etki rolüne sahip olabilir. (Örneğin mikroRNA)
Doğuştan gelenler - sonradan kazanılanlar

Genler, bir organizmanın işleyişiyle ilgili bütün enformasyonu içermekteyse de, çevre, nihai fenotipin belirlenmesinde önemli bir rol oynar. Genetik faktör ile çevre faktörü ikilemi, “doğuştan gelenler ile sonradan kazanılanlar” anlamında kullanılan, İngilizce “nature versus nurture” (kısaca, nature vs. nurture, doğa ve yetişme ikilemi) deyişiyle ifade edilir. Bir organizmanın fenotipi kalıtım ile çevrenin etkileşimine bağlıdır. “Isıya duyarlı mutasyonlar” olgusu bu duruma örnek olarak gösterilebilir. Genellikle, bir protein dizisi içinde değişen bir amino asit, onun davranışını ve diğer moleküllerle etkileşimini değiştirmez; fakat yapının istikrarını bozar. Yüksek sıcaklıkta moleküller daha hızlı hareket ettikleri ve birbirleriyle çarpıştıkları için, böylesi bir amino asit değişimi, proteinde yapısının bozulmasıyla (denatürasyon) ve işleyişinin zayıflamasıyla kendini gösteren bozukluklara yol açar. Düşük sıcaklıklı ortamlarda ise proteinin yapısı istikrarlı kalır ve işleyişi normal halde devam eder. Bu mutasyon türü siyam kedisinin kürkünde renk bakımından gözle görülür halde kendini gösterir: Pigment üretiminden sorumlu bir enzimdeki mutasyon, derideki yüksek sıcaklıklı bölgelerde yapısal istikrarının bozulmasına ve işleyişinin zayıflamasına yol açmaktayken bacak, kulak, kuyruk gibi daha soğuk bölgelerde protein, işleyişini zayıflatmadan sürdürür; böylece kedi, uç bölgeleri koyu renkli bir kürke sahip olur.[51]
Gen düzenlemesi
Bir organizmanın genomu binlerce gen içermekle birlikte, bu genlerin hepsinin de belirli bir anda aktif olmaları gerekmez. Bir gen, mRNA transkripsiyonu gerçekleştiğinde (ve proteine çevrildiğinde) “ifade olmuş” demektir. Genlerin ifadesini denetleyen birçok hücre yöntemi vardır. Mesela proteinler yalnızca hücre ihtiyaç duyduğunda üretilirler. Transkripsiyon faktörleri genin transkripsiyonunu ya teşvik etmek ya da engellemek suretiyle düzenleyen proteinlerdir.[52] Örneğin, Escherichia coli bakterisinin genomunda triptofan amino asitinin sentezi için gerekli bir seri gen vardır; fakat triptofanın hücrede kullanıma hazır hale gelmesinden sonra, bu genlere artık ihtiyaç kalmaz. Triptofanın varlığı genlerin faaliyetini doğrudan etkiler; triptofan molekülleri “triptofan represörü”ne (bir transkripsiyon faktörü) bağlanırlar, bağlanınca represörlerin yapısını öyle değiştirir ki, represörler genlere bağlanır. Triptofan represörü genlerin transkripsiyonu ve ifadesini durdurur, ve dolayısıyla, triptofan sentezi sürecinin “olumsuz geri beslemeli” (negative feedback) düzenlemesini sağlamış olur.[53]
Gen ifadesindeki farklılıklar, özellikle çok hücreli organizmalarda belirgindir, bu tip canlılarda hücrelerin hepsi aynı genomu içermelerine karşın, farklı gen kümelerinin ifadesinden kaynaklanan çok farklı yapı ve davranışlara sahiptirler. Çok hücreli bir organizmadaki tüm hücreler, tek bir hücreden türerler. Bu tek hücrenin farklı hücre tiplerine farklılaştığı süreç sırasında, dış ve hücreler arası sinyallere tepki verir, aşamalı olarak farklı gen ifade şekilleri kurarak farklı davranış tipleri oluşturur. Çok hücreli organizmalarda yapıların gelişiminden tek bir gen sorumlu değildir; bu farklı davranış tipleri birçok hücre arasındaki karmaşık etkileşimlerden doğar.
Ökaryotlarda kromatinde yapısal özellikler genlerin transkripsiyonunu etkiler. Bu özellikler “epigenetik”tir (üst-kalıtsal), çünkü etkileri DNA dizisinin üzerinde yer alır ve bir hücre kuşağından diğerine aktarılan kalıta haizdir. Epigenetik özelliklerden olayı, aynı ortamda oluşan farklı hücre tipleri çok farklı özelliklere sahip olabilirler.
Genetik değişim
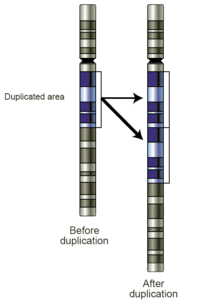
Mutasyonlar
DNA ikileşmesi süreci sırasında ikinci iplikçiğin polimerizasyonunda rastlantısal yanlışlıklar gerçekleşir. Mutasyon ya da değişinim adı verilen bu hatalar, özellikle bir genin protein kodlama dizisinde oluşmaları durumunda organizmanın fenotipi üzerinde güçlü bir etkide bulunabilirler. Fakat DNA polimeraz enziminin, hataları düzeltme yeteneği sayesinde bu hataların oranı son derece düşüktür; hata oranı, her 10-100 milyon bazda 1 hata olarak gözlemlenmiştir.[54][55] DNA’daki değişim oranını arttıran süreçlerin mutajenik olduğu söylenir. Mutajenik kimyasallar genellikle baz eşleşmesine müdahale ederek, DNA ikileşmesinde hatalara yol açarlar. Morötesi ışınım ise, DNA yapısına zarar vermek suretiyle mutasyonlara neden olur.[56] DNA’daki kimyasal zarar doğal yolla meydana gelmekteyse de, hücreler uyumsuzlukları ve bozulmaları tamir etmek üzere “DNA tamir” mekanizmalarını kullanırlar. Ancak, tamir bazen DNA’yı -dizisi bakımından- orijinal haline geri döndüremeyebilir.
Krosover ile kromozomal parça değişimi yapan ve genleri yeniden birleştiren (rekombine eden) organizmalarda mayoz bölünme esnasındaki hizalanma (iki kromozomdaki benzer dizilerin yan yana gelmesi) hataları da mutasyonlara neden olabilir.[57] Bu hatalar, benzer dizilerin neden oldukları, partner kromozomların hatalı hizalanması sonucu olması özellikle muhtemeldir; bu da genomlardaki bazı bölgeleri mutasyona daha eğilimli kılar. Bu hatalar DNA dizisinde büyük yapısal değişiklikler yaratır; kromozomda geniş bölgelerde duplikasyonlar (ikilenmeler), inversiyonlar (evirmeler), delesyonlar (çıkarmalar) veya farklı kromozomlar arasında parçaların kazara aktarılması (translokasyon) sözkonusu olabilir.
Doğal seçilim ve evrim
Mutasyonlar farklı genotipli organizmaların ortaya çıkmasına neden olur ve bu farklılıklar da farklı fenotiplerin oluşmasıyla sonuçlanır. Birçok mutasyonun organizmanın fenotipi, sağlığı ve (doğal seçilimle ilgili) üreme uyumu (İng. fitness) üzerinde az bir etkisi vardır. Etkisi olan mutasyonlar genelde zararlıdırlar ama bazen, organizmanın içinde bulunduğu çevre koşulları bağlamında yararlı denebilecek mutasyonlar da olur.
Popülasyon genetiği popülasyonlardaki bu genetik farklılıkların kaynaklarını, dağılımlarını ve bu dağılımların zamanla nasıl değiştiğini araştıran bir genetik altdalıdır.[58] Bir alelin bir popülasyondaki sıklığı doğal seçilimle etkilenebilir; belirli bir aleli taşıyan bireylerin hayatta kalma ve üremesindeki yüksek oran, o alelin zamanla o popülasyonda daha sık olmasına neden olabilir.[59] Aynı zamanda, “genetik sürüklenme” denilen, şans faktörünün etkisiyle, yani olayların tesadüfi akışıyla da, allel sıklığında değişimler olabilir.[60] Genetik sürüklenme bir popülasyonun gen havuzunda, doğal seçilimden farklı olarak, uygun genlerin seçilmesi gibi bir yönlendirmeyle değil de, tamamen rastlantı eseri sayılan, kuşaktan kuşağa ortaya çıkan değişiklikler şeklinde tanımlanır.
Organizmaların genomları, birçok kuşak boyunca, evrim denilen olgu ile sonuçlanmak üzere, değişebilirler. Mutasyonlar ve mutasyonların yararlı olanları için olan seçilim sonucunda, bir canlı türün çevresine daha uyumlu biçimlere dönüşerek evrimine neden olabilir. Bu sürece adaptasyon denir.[61] Yeni türler, türleşme denilen süreçle oluşur. Türleşme genellikle, farklı popülasyonların coğrafi olarak ayrı düşmelerinin neden olduğu genetik farklılaşmadan ortaya çıkar.[62]
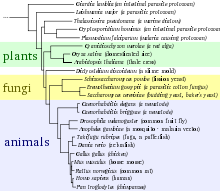
Evrim esnasında DNA dizileri birbirinden uzaklaştığı ve değiştiği için, diziler arasındaki bu farklılıklar, aralarındaki evrimsel uzaklığı hesaplamada bir “moleküler saat” gibi kullanılabilir.[63] Genetik kıyaslamalar genellikle, türler arasındaki evrimsel akrabalığı nitelemede en doğru yöntem olarak kabul edilir, bu yöntem, fenotipik kıyaslamalarla edinilmiş bazı yanıltıcı değerlendirmeleri de düzeltir. Türler arasındaki evrimsel uzaklıklar “evrim ağacı” ya da “filogenetik ağaç” denilen şemalarla temsil edilir, bu şemalarla türlerin ortak bir atadan inişini ve zaman boyunca türlerin birbirinden uzaklaşmalarını gösterir. Ancak, bu ağaç şemaları türler arasındaki yatay gen transferi olaylarını gösteremez.
Araştırma ve teknoloji
.jpg)
Model organizmalar
Genetikçiler başlangıçta genetiği geniş bir organizma yelpazesi üzerinde çalışmışlarsa da, sonraları araştırmacılar organizmaların bir altkümesi üzerinde özelleşmeye başlamıştır. Belli bir organizma hakkında önemli miktarda araştırma yapılmış olması yeni araştırmacıların da aynı organizmayı daha derinlemesine icelemeye teşvik etmiştir. Böylece birkaç model organizma günümüzdeki genetik araştırmaların önemli bir kısmı için temel oluşturmuştur.[64] Model organizmalar genetiğindeki başlıca araştırma konuları, gen düzenlemesi, morfogeneze ilişkin gelişim genleri ve kanserdir.
Model organizmalar kısmen kullanımlarının pratik olması nedeniyle seçilmiştir; kısa üretim süreleri, genetik manipülasyonun kolay olması bazı organizmaların genetik araştırmalarda popüler olmasına neden olmuştur. Yaygın olarak kullanılan model organizmalar arasında, bağırsak bakterisi Escherichia coli, turpgiller familyasından Arabidopsis thaliana bitkisi, bir maya türü olan Saccharomyces cerevisiae, iplik kurdu Caenorhabditis elegans , yaygın meyve sineği Drosophila melanogaster ve ev faresi Mus musculus sayılabilir.
Farklı araştırma alanları
Genetik bilimindeki gelişmelerin yanı sıra, araştırmaların gitgide farklı alanlarda özelleşmeye başlaması bu bilim dalının altdallarının oluşmasına neden olmuştur. Genetiğin altdallarından bazıları şunlardır:
- Evrimsel gelişim genetiği (Génétique évolutive du développement): Döllenmiş tekhücreli yumurta aşamasından başlayarak organizmanın oluşmasındaki tüm moleküler etkenleri ve dolayısıyla onları kodlayan genleri inceler. Yoğun olarak, özellikle iki taraflı simetri düzenlenmesiyle ve basit bir biyolojik sistemden (tekhücreliler, ışınsal simetri) karmaşık bir organizmaya (çokhücreli, genellikle metamerize ve özelleşmiş organlar halinde yapılaşmış organizmalar) geçişi sağlayan mekanizmalarla ilgilenir. Organizmanın oluşum mekanizmalarını incelemek için model organizma türleri (Drosophila , yuvarlak solucanlar, zebra balığı, tavuk vs.) kullanır. Fransızca'da evrimsel gelişim genetiği adıyla bilinen bu dal, İngilizce'de evrimsel gelişim biyolojisi olarak bilinir.
- Medikal genetik
- Genomik: İnsan genomunun (kromozomlarda yapılanmış üç milyar baz çiftinin, DNA bütününün) yapısını, bileşimini ve evrimini inceler ve DNA’da biyolojik bir anlamı olabilecek birimleri (genler, çevrilmeyen transkripsiyon birimleri, mikroRNA’lar, düzenleme üniteleri, transkripsiyon faktörleri olan promotörler, CNG alfa ve beta kanalları vs.) tanımlamaya çalışır.
- Kantitatif genetik : Genetik bileşenleri, niceliksel özelliklerin (boy, tüy rengi, büyüme hızı vs.) varyasyonunu (değişme, çeşitlenme) ve kalıtsallıklarını açıklayarak inceler.
- Evrim genetiği : Türlerin genomlarında doğal seçilimin izlerini inceler ve türlerin değişen çevrelerde (ortamlarda) hayatta kalmasında ve adaptasyonunda baş rolü oynayan genleri tanımlamaya çalışır.
- Popülasyon genetiği: Popülasyonların ve türlerin çeşitliliğini etkileyen güçleri (ve etki ya da sonuçlarını) matematiksel ve istatistikî yöntemler geliştirerek inceler. Bir başka deyişle popülasyonlardaki fertlerin benzerlik ve farklılıklarının kaynaklarını araştıran bir genetik altdalıdır. Dört ana madde üzerinden yola çıkarak araştırmalar yapar: Bunlar doğal seçilim, gen havuzu, mutasyonlar ve gen devamlılığıdır.
- Moleküler genetik: Canlıların kalıtım materyali olan genlerin yapılarını ve işlevlerini moleküler düzeyde inceleyen bir genetik altdalıdır. Moleküler genetik, moleküler biyolojinin ve genetiğin yöntemlerini kullanarak çalışır.
- Ekolojik genetik: Genetik çalışmaları ekolojik alanda sürdüren bir genetik altdalıdır. Ekolojik genetik, canlıların oluşturduğu popülasyonları "popülasyon genetiği" ile yakından ilişkili olarak araştırır.
Medikal genetik araştırmaları
Medikal genetik, genetik çeşitliliğin, insan sağlığı ve hastalıklarıyla ilişkilerini araştırmaktadır.[65] Bir hastalığa neden olabilecek bilinmeyen bir gen araştırıldığında, araştırmacılar, hastalıkla ilgili genomun konumunu saptamada genellikle “genetik bağlantı” ve genetik soyağacı çizelgesinden yararlanırlar. Popülasyon düzeyindeki araştırmalarda, araştırmacılar genomdaki, hastalıklarla ilgili genlerin konumlarını saptamada “Mendelci rastgeleleştirme” yönteminden yararlanmaktadır; bu teknik bilhassa, yalnızca tek bir genle kesin olarak belirlenemeyen, birkaç gene ilişkin (çok genli) özelliklerde yararlı olmaktadır.[66] Hastalık geni olabilecek herhangi bir gen aday olarak saptanınca, artık sonraki araştırmalar genellikle, bu genin bir model organizmadaki dengi olan gen (ortolog gen) üzerinde yapılır. Genotipleme teknikleri, kalıtımsal hastalık çalışmalarının yanı sıra, genotipin ilaca cevabı nasıl etkilediğini araştıran farmakogenetik alanının gelişmesini de sağlamıştır.[67]
Kanser kuşaktan kuşağa kalıtım yoluyla geçen bir hastalık olmasa da, günümüzde genetik bir hastalık olarak ele alınmaktadır.[68] Kanserin vücuttaki gelişim süreci çeşitli olayların bir araya gelmesiyle oluşmaktadır. Bazen vücuttaki hücreler bölünürken mutasyonlar olur. Bu hücrelerdeki mutasyonlar bir çocuğa aktarılmasa da, hücrelerin davranışını etkileyebilmekte ve kimi zaman onların büyümelerine ve daha hızlı bölünmelerine neden olmaktadırlar. Hücrelerin bu anormal ve uygunsuz bölünmelerini engelleyen mekanizmalar vardır; uygunsuz bölünmekte olan hücrelerin ölmesi için sinyaller yolanır. Ama bazen başka mutasyonlar çoğalan hücrelerin bu sinyallere uymamasına neden olabilir. Vücutta, bir çeşit dahili bir doğal seçilim süreci meydana gelir; hücrenin bölünmeye devamını sağlayan mutasyonlara hücrelerde birikir, sonunda bir kanser tümörü meydana gelir. Tümör büyüyüp gelişerek vücudun çeşitli dokularını istila eder.
. E. coli rekombinant DNA teknolojisinde sıkça kullanılır.
Araştırma teknikleri
Günümüzde DNA, laboratuvarda birçok bakımdan istenildiği gibi değiştirilebilmektedir. Laboratuvar çalışmalarında kullanılan restriksiyon enzimleri DNA’yı belli dizilerde keserek arzu edilen parçaları üretmek için kullanılır.[69] Ligasyon enzimleri ise, elde edilen bu parçaları yeniden birleştirme, yani birbirine bağlama olanağı sağlamaktadır ve böylece, araştırmacılar, farklı kaynaklardan (biyolojik türlerden) alınan DNA parçalarını birleştirerek “rekombinant DNA” yaratabilmektedirler. Genellikle “genetik yapısı değiştirilmiş organizmalar”la (İngilizce kısaltmasıyla GMO) ilgili çalışmalarda yararlanılan rekombinant DNA bilhassa, plazmidler (üzerlerinde birkaç gen bulunan dairesel DNA parçaları) bağlamında kullanılmaktadır. Bakterilerin içine plazmidlerin sokulması ve bu bakterilerin “agar” tabaklarında (bakteri hücrelerinin klonlarını izole etmek için) büyütülmesiyle araştırmacılar, eklenen DNA parçalarını klonal olarak çoğaltabilmektedirler ki bu, moleküler klonlama olarak bilinen bir işlemdir. (Klonlama terimi, aynı zamanda çeşitli teknikler kullanarak klonal organizmalar yaratmak için de kullanılır.)
DNA aynı zamanda polimeraz zincir tepkimesi (PCR) denilen bir süreç kullanılarak da çoğaltılabilir.[70] PCR, özel kısa DNA dizileri kullanılarak, DNA’nın hedef seçilen bir bölgesini izole edebilir ve onu aşırı derecede büyütebilir. DNA’nın son derece küçük parçalarını aşırı ölçüde çoğaltabildiğinden, PCR genellikle spesifik DNA dizilerinin varlığını saptamakta kullanılır.
DNA dizilemesi ve genomik

Genetik çalışmalarında geliştirilmiş en temel teknolojilerden biri olan DNA dizilemesi araştırmacılara DNA parçalarındaki nükleotit dizisini belirleme olanağı sağlamaktadır. 1977’de Frederick Sanger ve çalışma arkadaşlarınca geliştirilen bir DNA dizileme yöntemi (zincir sonlandırma dizilemesi) DNA parçalarını dizilemede artık rutin bir yöntem olarak kullanılmaktadır.[71] Bu teknoloji sayesinde araştırmacılar, birçok insan hastalığıyla ilgili moleküler dizileri inceleme olanağına kavuşmuşlardır.
DNA dizilemesi ucuzlaştıkça ve bilgisayarların da yardımıyla araştırmacılar, birçok organizmanın genomunu dizilemişlerdir. Bunu yapmak için dizilenmiş DNA parçaları, dizilerinin aynı olduğu bölgeleri çakıştırılarak, daha büyük bölgelerin dizileri belirlenir (genom inşası süreci) dizilemişlerdir.[72] Bu teknolojiler, insan genomu için de kullanılmış, insan genomunun dizileme projesi 2003 yılında tamamlanmıştır.[23] Yeni yüksek hacimli dizileme teknolojileri DNA dizileme maliyetini hızla düşürmektedir, çoğu araştırmacı bir insan genomunun dizilenme maliyetinin yakın gelecekte bin dolara inmesini beklemektedir.
DNA dizileme yöntemleriyle belirlemeler sonucunda edinilen, işe yarar dizilemelerin miktarının gitgide artması, organizmaların genom bütünlerindeki araştırmalarda hesaplama aletleri ve analiz örnekleri kullanan, genomik adlı araştırma alanını doğurmuştur. Genomik aynı zamanda, biyoenformatik bilimsel disiplininin bir altalanı olarak da kabul edilebilir.
Dış bağlantılar
- Genetik
- Genetics-1
- Genetics-2
- Genetics Education Center
- Free database on the human genetics
- Database of the genomes sequences, as well as the projects in progress
- Discovered in 2007, the zones of instability of the genome in the mouse.
- Yapay DNA
- İnsan klonlama kararı
- Hayvanlardan organ nakline denetim
- İnsan embriyosunda genetik değişiklik yapıldı
- Yapay döllenme
- Biological weapons,genetics and social analysis
- Genetic engineering and biological weapons
- Genetic Bullets - Ethnically Specific Bioweapons
- Biological warfare, bioterrorism, biodefence and the biological and toxin weapons convention
- Ethnic Specific Weapons
- (Fransızca) Glossaire de la biotechnologie de la FAO
- (Fransızca) «Il était une fois ... l'ADN»: un site éducatif sur les bases de la génétique classique et moléculaire
- (Fransızca) Du gène à la compréhension de la molécule qu'elle code dans l'organisme. Une vidéo éducative en partenariat avec l'Institut Pasteur
- (Fransızca) Un site présentant les aspects éthiques liés à la question de la manipulation génétique des plantes et des animaux. Les aspects "sciences" et "droits" sont en construction.
- (Fransızca) Génétique dans l'élevage chiens de race: consanguinité, retrempe, gènes codant la couleur de la robe, la longueur de la queue.
Ayrıca bakınız
- Mendel
- Mendel genetiği
- Genler ve Alleller
- Monohibrit çaprazlama
- Dihibrit çaprazlama
- Genotip
- Fenotip
- Mutasyonlar
- Eşey saptanması
- Bölünme
- Kromozomlar
- Klonlama
- Gen aktarımı
- Genetik tanı merkezi
| ||||||||||
| ||||||||||||||
Kaynakça
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K, and Walter P (2002). Molecular Biology of the Cell, 4th edition. ISBN 0-8153-3218-1.
- Griffiths AJF, Miller JH, Suzuki DT, Lewontin RC, and Gelbart WM (2000). An Introduction to Genetic Analysis. New York: W.H. Freeman and Company. ISBN 0-7167-3520-2.
- Hartl D, Jones E (2005). Genetics: Analysis of Genes and Genomes, 6th edition. Jones & Bartlett. ISBN 0-7637-1511-5.
- Lodish H, Berk A, Zipursky LS, Matsudaira P, Baltimore D, and Darnell J (2000). Molecular Cell Biology, 4th edition. ISBN 0-7167-3136-3.
- Salt, Alparslan. Geleceğin Felaketleri, Ruh ve Madde Yayınları, İstanbul, 1997
- Asimov,I. İnsanlığın Geleceği, Cep Kitapları,İstanbul,1984
- Naisbitt,J. ve Aburden,P. Megatrends 2000,Form Yayınları,1990
- Cumhuriyet gazetesi, Bilim ve Teknik ilave dergileri, 1992-1995 yılları arasındaki sayılar.
- İngilizce ve Fransızca Wikipedia'daki 9 Mayıs 2008 tarihli genetik maddesi.
Notlar ve referanslar
- 1 2 "genetik." Güncel Türkçe Sözlük. Türk Dil Kurumu. Erişim: 2 Ağustos 2012
- ↑ Griffiths et al. (2000), Chapter 1 (Genetics and the Organism): Introduction
- ↑ Hartl D, Jones E (2005)
- ↑ "genetic." Online Etimology Dictionary. Erişim: 2 Ağustos 2012.
- ↑ Weiling F (1991). "Historical study: Johann Gregor Mendel 1822–1884". American Journal of Medical Genetics 40 (1): 1–25; discussion 26. PMID 1887835.
- ↑ Augustinians
- ↑ Mendel, GJ (1866). "Versuche über Pflanzen-Hybriden]". Verhandlungen des naturforschenden Vereins Brünn 4: 3–47. (in English in 1901, J. R. Hortic. Soc. 26: 1–32) English translation available online
- ↑ "Gregor Mendel The Beginning of Biomathematics". http://web.archive.org/web/20080117233318/http://mendel.imp.ac.at/mendeljsp/biography/biography.jsp. Erişim tarihi: 21-12-2008.."Mendel's Methods and Statistics". 11 Mart 2015 tarihinde kaynağından arşivlendi. http://web.archive.org/web/20150311193751/http://www.biology.ewu.edu:80/aHerr/Genetics/Bio310/Pages/ch3pages/ch3page.html. Erişim tarihi: 21-12-2008.."Statistical methods in Bioinformatics". http://webcache.googleusercontent.com/search?q=cache:L607ZQc9SK4J:www.math.ucsd.edu/~gptesler/283/links.html+Mendel+statistic+method&hl=tr&ct=clnk&cd=31&gl=tr. Erişim tarihi: 21-12-2008.."Physiology or Medicine 1933". http://webcache.googleusercontent.com/search?q=cache:5O0pYT-eq8MJ:nobelprize.org/nobel_prizes/medicine/laureates/1933/press.html+Mendel+statistic+method&hl=tr&ct=clnk&cd=22&gl=tr. Erişim tarihi: 21-12-2008.."The Mathematics of Inheritance". http://webcache.googleusercontent.com/search?q=cache:Ikwww8PbcMQJ:www.mendel-museum.org/eng/1online/room2.htm+Mendel+statistic+method&hl=tr&ct=clnk&cd=25&gl=tr. Erişim tarihi: 21-12-2008.
- ↑ Genetics, n., Oxford English Dictionary, 3rd ed.
- ↑ Bateson W. Letter from William Bateson to Alan Sedgwick in 1905. The John Innes Centre. Retrieved on 2008-03-15.
- ↑ Bateson, W (1907). "The Progress of Genetic Research". Wilks, W (editor) Report of the Third 1906 International Conference on Genetics: Hybridization (the cross-breeding of genera or species), the cross-breeding of varieties, and general plant breeding, London: Royal Horticultural Society.
- ↑ genetic, adj., Oxford English Dictionary, 3rd ed.
- ↑ Moore JA (1983). "Thomas Hunt Morgan—The Geneticist". American Zoologist 23 (4): 855–865.
- ↑ Sturtevant AH (1913). "The linear arrangement of six sex-linked factors in Drosophila, as shown by their mode of association". Journal of Experimental Biology 14: 43–59. pdf from Electronic Scholarly Publishing
- ↑ Avery OT, MacLeod CM, and McCarty M (1944). "Studies on the Chemical Nature of the Substance Inducing Transformation of Pneumococcal Types: Induction of Transformation by a Desoxyribonucleic Acid Fraction Isolated from Pneumococcus Type III". Journal of Experimental Medicine 79 (1): 137–158. 35th anniversary reprint available
- ↑ Hershey AD, Chase M (1952). "Independent functions of viral protein and nucleic acid in growth of bacteriophage". The Journal of General Physiology 36: 39–56. doi:
- ↑ Judson, Horace (1979). The Eighth Day of Creation: Makers of the Revolution in Biology. Cold Spring Harbor Laboratory Press, 51–169. ISBN 0-87969-477-7.
- ↑ Watson JD, Crick FHC (1953). "Molecular structure of Nucleic Acids: A Structure for Deoxyribose Nucleic Acid" (PDF). Nature 171 (4356): 737–738.
- ↑ Watson JD, Crick FHC (1953). "Genetical Implications of the Structure of Deoxyribonucleic Acid" (PDF). Nature 171 (4361): 964–967.
- ↑ History of genetic code.Discoverer of the genetic code.Crack the code. Genetic code.
- ↑ Sanger F, Nicklen S, and Coulson AR (1977). "DNA sequencing with chain-terminating inhibitors". Nature 74 (12): 5463–5467. doi:10.1073/pnas.74.12.5463.
- ↑ Saiki RK, Scharf S, Faloona F, Mullis KB, Horn GT, Erlich HA, Arnheim N (1985). "Enzymatic Amplification of β-Globin Genomic Sequences and Restriction Site Analysis for Diagnosis of Sickle Cell Anemia". Science 230 (4732): 1350–1354. PMID 2999980.
- 1 2 Human Genome Project Information. Human Genome Project. Retrieved on 2008-03-15.
- ↑ Griffiths et al. (2000), Chapter 2 (Patterns of Inheritance): Introduction
- ↑ Mendel, GJ (1866). "Versuche über Pflanzen-Hybriden". Verhandlungen des naturforschenden Vereins Brünn 4: 3–47. (in English in 1901, J. R. Hortic. Soc. 26: 1–32) English translation available online
- ↑ Griffiths et al. (2000), Chapter 2 (Patterns of Inheritance): Mendel's experiments
- ↑ Çaprazlama (cross-breed, cross-breeding, cross-fertilize). Ayrıca bkz. Monohibrit çaprazlama
- 1 2 3 Griffiths et al. (2000), Chapter 3 (Chromosomal Basis of Heredity): Mendelian genetics in eukaryotic life cycles
- ↑ Griffiths et al. (2000), Chapter 4 (Gene Interaction): Interactions between the alleles of one gene
- ↑ Richard W. Cheney.Genetic Notation. Retrieved on 2008-03-18.
- ↑ Griffiths et al. (2000), Chapter 2 (Patterns of Inheritance): Human Genetics
- ↑ Griffiths et al. (2000), Chapter 4 (Gene Interaction): Gene interaction and modified dihybrid ratios
- ↑ Mayeux R (2005). "Mapping the new frontier: complex genetic disorders". The Journal of Clinical Investigation 115 (6): 1404–1407. doi:10.1172/JCI25421. PMID 15931374.
- ↑ Griffiths et al. (2000), Chapter 25 (Quantitative Genetics): Quantifying heritability
- ↑ Luke A, Guo X, Adeyemo AA, Wilks R, Forrester T, Lowe W Jr, Comuzzie AG, Martin LJ, Zhu X, Rotimi CN, Cooper RS (2001). "Heritability of obesity-related traits among Nigerians, Jamaicans and US black people". Int J Obes Relat Metab Disord 25 (7): 1034–1041. Abstract from NCBI
- ↑ Pearson H (2006). "Genetics: what is a gene?". Nature 441 (7092): 398–401. PMID 16724031.
- ↑ Prescott, L (1993). Microbiology. Wm. C. Brown Publishers. 0-697-01372-3.
- ↑ Griffiths et al. (2000), Chapter 8 (The Structure and Replication of DNA): Mechanism of DNA Replication
- ↑ Gregory SG et al. (2006). "The DNA sequence and biological annotation of human chromosome 1". Nature 441. doi: 10.1038/nature04727. free full text available
- ↑ Alberts et al. (2002), DNA and chromosomes: Chromosomal DNA and Its Packaging in the Chromatin Fiber
- ↑ Griffiths et al. (2000), Chapter 2 (Patterns of Inheritance): Sex chromosomes and sex-linked inheritance
- ↑ Griffiths et al. (2000), Chapter 7 (Gene Transfer in Bacteria and Their Viruses): Bacterial conjugation
- ↑ Griffiths et al. (2000), Chapter 7 (Gene Transfer in Bacteria and Their Viruses): Bacterial transformation
- ↑ Griffiths et al. (2000), Chapter 5 (Basic Eukaryotic Chromosome Mapping): Nature of crossing-over
- ↑ Griffiths et al. (2000), Chapter 5 (Basic Eukaryotic Chromosome Mapping): Linkage maps
- ↑ Gen ifadesi
- ↑ Berg JM, Tymoczko JL, Stryer L, Clarke ND (2002). Biochemistry, 5th edition, New York: W. H. Freeman and Company. I. 5. DNA, RNA, and the Flow of Genetic Information: Amino Acids Are Encoded by Groups of Three Bases Starting from a Fixed Point
- ↑ Crick, F (1970): Central Dogma of Molecular Biology (PDF). Nature 227, 561–563. PMID 4913914
- ↑ Alberts et al. (2002), Proteins: The Shape and Structure of Proteins .Alberts et al. (2002), Proteins: Protein Function
- ↑ How Does Sickle Cell Cause Disease?. Brigham and Women's Hospital: Information Center for Sickle Cell and Thalassemic Disorders (2002-04-11). Retrieved on 2007-07-23.
- ↑ Imes DL, Geary LA, Grahn RA, Lyons LA (2006). "Albinism in the domestic cat (Felis catus) is associated with a tyrosinase (TYR) mutation" (Short Communication). Animal Genetics 37 (2): 175. doi:10.1111/j.1365-2052.2005.01409.x. Retrieved on 2006-05-29.
- ↑ Brivanlou AH, Darnell JE Jr (2002). "Signal transduction and the control of gene expression". Science 295 (5556): 813–818.PMID 11823631.
- ↑ Alberts et al. (2002),Control of Gene Expression - The Tryptophan Repressor Is a Simple Switch That Turns Genes On and Off in Bacteria
- ↑ Griffiths et al. (2000),Chapter 16 (Mechanisms of Gene Mutation): Spontaneous mutations
- ↑ Kunkel TA (2004). "DNA Replication Fidelity". Journal of Biological Chemistry 279 (17): 16895–16898. doi:10.1038/sj.emboj.7600158.
- ↑ Griffiths et al. (2000),Chapter 16 (Mechanisms of Gene Mutation): Induced mutations
- ↑ Griffiths et al. (2000),Chapter 17 (Chromosome Mutation I: Changes in Chromosome Structure): Introduction
- ↑ Griffiths et al. (2000),Chapter 24 (Population Genetics): Variation and its modulation
- ↑ Griffiths et al. (2000),Chapter 24 (Population Genetics): Selection
- ↑ Griffiths et al. (2000),Chapter 24 (Population Genetics): Random events
- ↑ Darwin, Charles (1859).On the Origin of Species, 1st, John Murray, 1. . Related earlier ideas were acknowledged in Darwin, Charles (1861).On the Origin of Species, 3rd, John Murray, xiii.
- ↑ Gavrilets S (2003). "Perspective: models of speciation: what have we learned in 40 years?". Evolution 57 (10): 2197–2215. doi:10.1554/02-727.PMID 14628909.
- ↑ Wolf YI, Rogozin IB, Grishin NV, Koonin EV (2002). "Genome trees and the tree of life". Trends Genet. 18 (9): 472–479.PMID 12175808.
- ↑ The Use of Model Organisms in Instruction. University of Wisconsin: Wisconsin Outreach Research Modules. Retrieved on 2008-03-15.
- ↑ NCBI: Genes and Disease. NIH: National Center for Biotechnology Information. Retrieved on 2008-03-15.
- ↑ Davey Smith, G; Ebrahim, S (2003)."‘Mendelian randomization’: can genetic epidemiology contribute to understanding environmental determinants of disease?". International Journal of Epidemiology 32: 1–22.
- ↑ Pharmacogenetics Fact Sheet. NIH: National Institute of General Medical Sciences. Retrieved on 2008-03-15.
- ↑ Strachan T, Read AP (1999). Human Molecular Genetics 2, second edition, John Wiley & Sons Inc.. Chapter 18: Cancer Genetics
- ↑ Lodish et al. (2000),Chapter 7: 7.1. DNA Cloning with Plasmid Vectors
- ↑ Lodish et al. (2000),Chapter 7: 7.7. Polymerase Chain Reaction: An Alternative to Cloning
- ↑ Brown TA (2002). Genomes 2, 2nd edition. ISBN ISBN 1-85996-228-9. Section 2, Chapter 6: 6.1. The Methodology for DNA Sequencing
- ↑ Brown (2002),Section 2, Chapter 6: 6.2. Assembly of a Contiguous DNA Sequence